
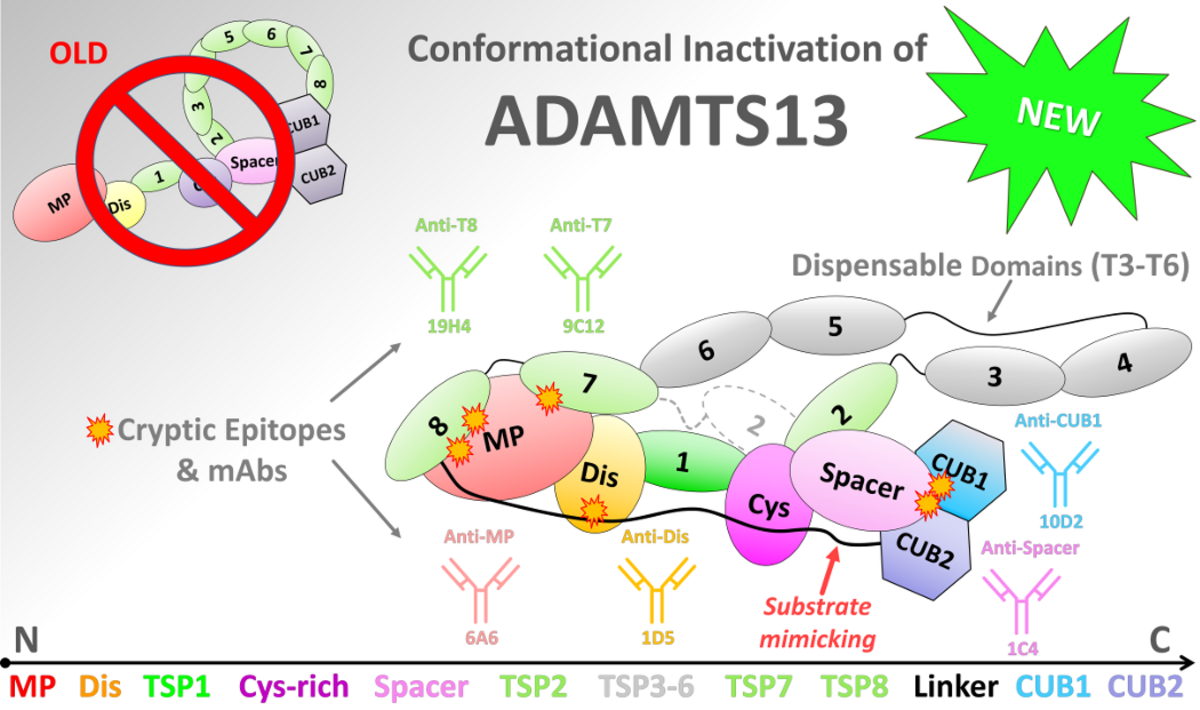

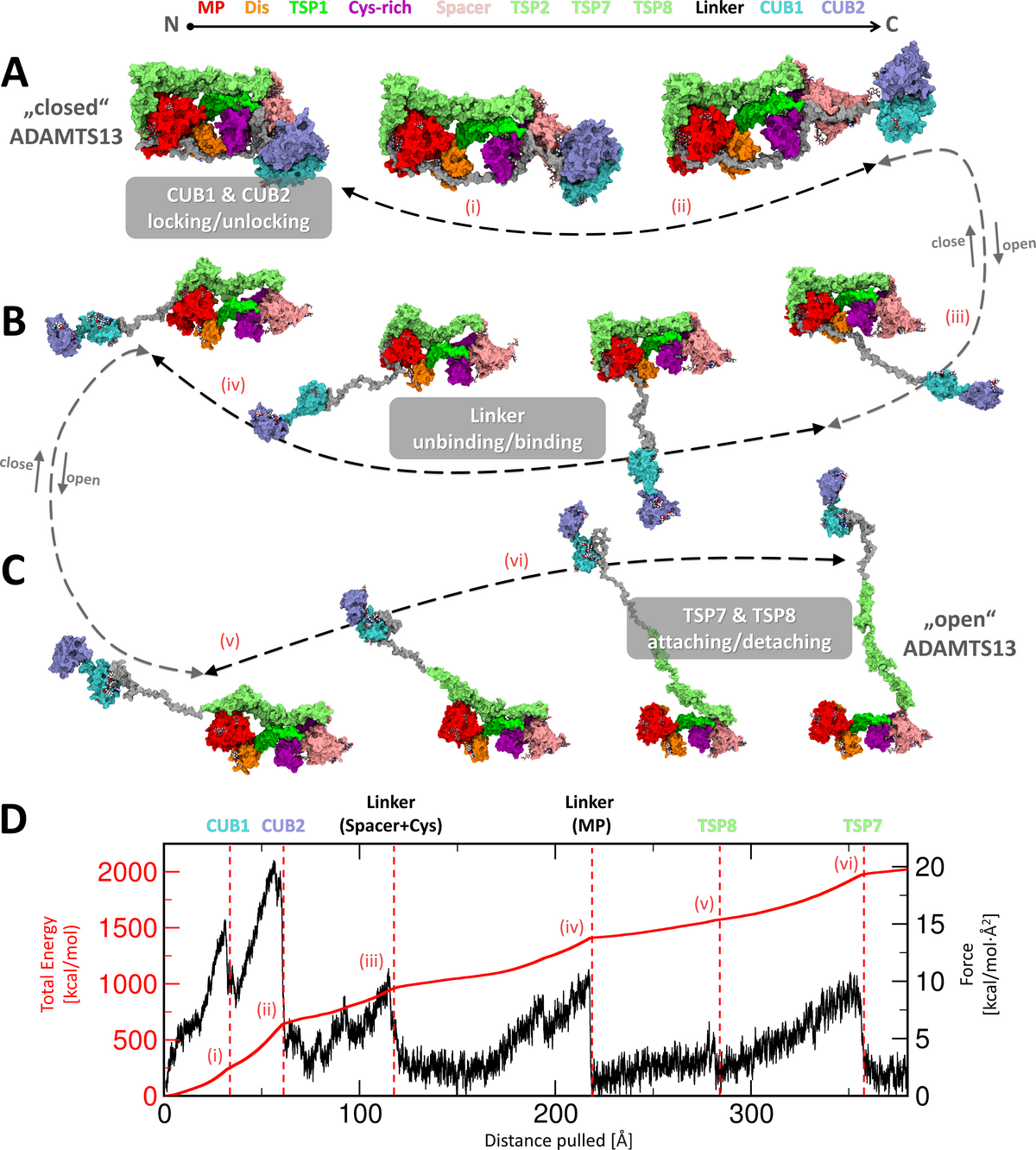

We have uncovered a previously enigmatic long-range regulation mechanism of ADAMTS13, a key enzyme that prevents dangerous blood clot formation in the circulation. Our study, titled “Wrapping it Up: Structural Basis of ADAMTS13 Global Latency”, now published in the Journal of Thrombosis and Haemostasis, provides the most detailed structural model to date explaining how ADAMTS13 maintains its inactive “closed” state.
Using large-scale molecular dynamics simulations together with biochemical experiments, we discovered that ADAMTS13 is controlled through a coordinated network of interactions across multiple domains of the enzyme. Central to the mechanism is the so-called “L3 Linker,” which acts as a pseudosubstrate and blocks critical substrate-binding regions. We describe this architecture as a “molecular zipper” that stabilizes the enzyme in its inactive state while still allowing rapid activation when needed.
The computational work was enabled by TIGER2hPE, a custom-developed enhanced-sampling simulation algorithm developed at the University of Greifswald. Even with this approach, the project required several cumulative years of computation time on the Göttingen high-performance computing infrastructure, part of NHR-NORD.
The study was coordinated by Norman Geist under the supervision of Mihaela Delcea in collaboration with Quintijn Bonnez and Karen Vanhoorelbeke at KU Leuven.
Beyond advancing the fundamental understanding of blood coagulation, our findings may support future precision therapies for thrombotic diseases such as thrombotic thrombocytopenic purpura (TTP).
Fulltext available at https://www.sciencedirect.com/science/article/pii/S1538783626003272