Norman's paper entitled "Wrapping it Up: Structural Basis of ADAMTS13 Global Latency" is published
Graphical Abstract
Opening-/Closing animation
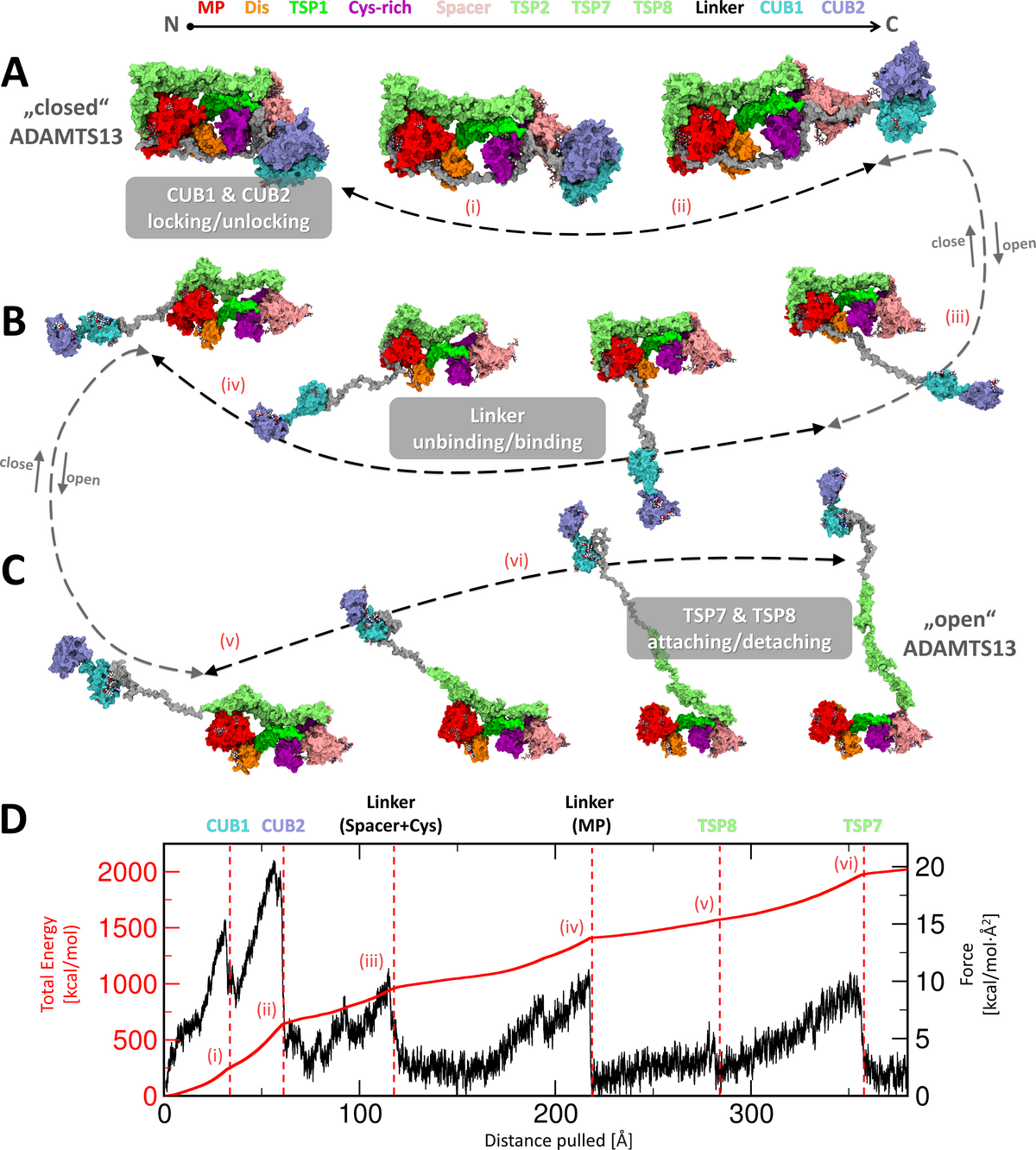
Figure 6: Conformational activation and inactivation mechanisms during global latency of ADAMTS13.
Domain colors and sequential order are indicated at the top. (A) (left) Our final closed model of ADAMTS13-Del3To6
shows both terminal CUB domains attached to Spacer, the Linker region covers the substrate-binding sites on the MP,
Dis, and Cys-rich domains, while TSP7 and TSP8 bound near the MP domain. Pathway for conformational
activation/inactivation illustrated with arrows. A movie illustrating this process intuitively is included in the
provided Zenodo archive. During activation, the CUB1 and CUB2 domains detach from the Spacer module. Roman
numerals (i-vi) highlight major rupture events observed during forced unfolding simulation (Sim6 ). (B) The Linker
region further disengages, exposing substrate-binding exosites on the Cys-rich, Dis, and MP domains. (C) The TSP8
and TSP7 domains detach from the MP domain, completing the conformational activation of ADAMTS13. (D)
Exemplary force and accumulated energy profiles derived from the forced unfolding simulation. (Structural
snapshots relaxed with MD for natural appearance)
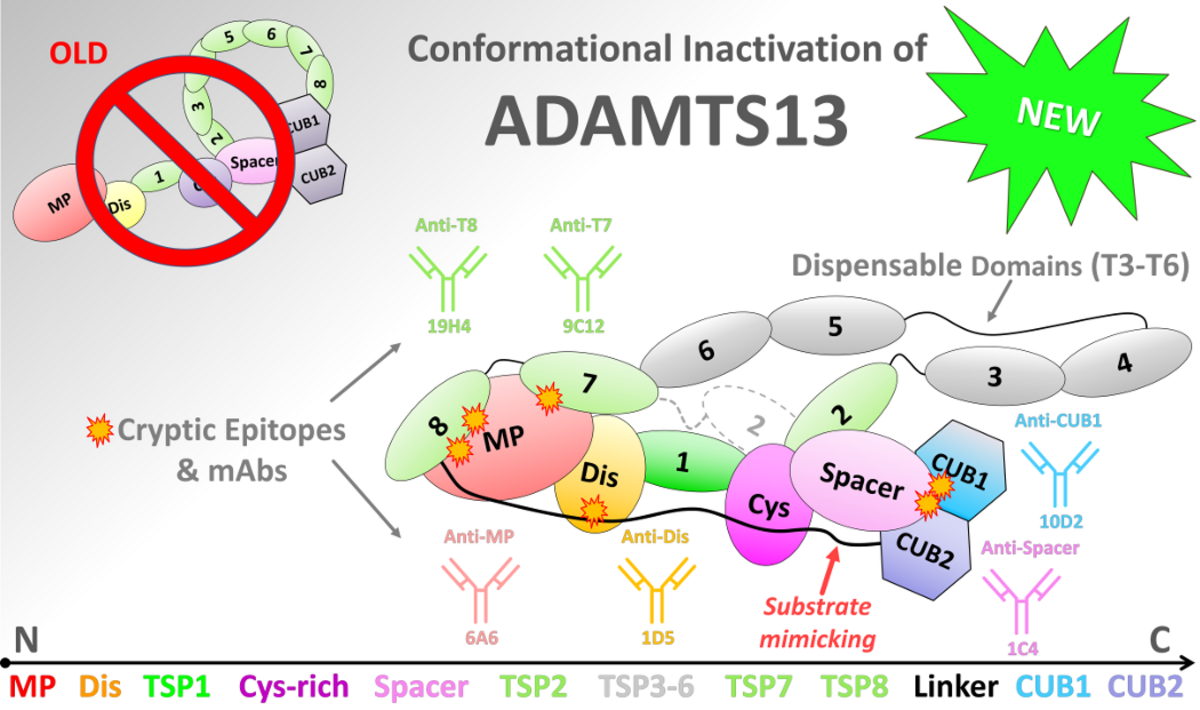
Figure 7: Conformation testing of full-length ADAMTS13 under environmental perturbations using ELISA.
(top) Schematic depiction of updated model on conformationally latent ADAMTS13. Domain colors and sequential order
are indicated at the top. The CUB domains rest at Spacer. Other distal domains are wrapped around the substrate
binding domains and shield off various cryptic epitopes for which mAbs were recently identified. The TSP3-6 repeats are
dispensable. The Linker is a pseudosubstrate, acting like VWF-A2, binding to substrate exosites of MP, Dis, and
Cys-rich modules. TSP7 and TSP8 rest at the MP domain. (A) Closed plasma ADAMTS13 incubated at pH 7.2
(filled bars) or pH 6.0 (open bars). In ELISA, binding of mAbs 6A6, 1D5, 1C4, 9C12, 19H4 and 10D2, directed
against the MP, Dis, Spacer, TSP7, TSP8 and CUB1 domains, was assessed. All mAbs potently bind to open, but
not to closed ADAMTS13. (B) Closed plasma ADAMTS13 was incubated in the absence (filled bars) or presence
(open bars) of EDTA. In ELISA, binding of mAbs was assessed as previously. Chelation of Zn2+ and Ca2+ ions
destabilized the structure of the MP domain resulting in abolished binding for mAbs 3H9 and 6A6, leading to partial
unwrapping of the expected domains. (relative binding (mean
± one standard deviation) to open ADAMTS13 mediated by mAb 18H10. Significance tested using two-way ANOVA,
including multiple comparisons with Bonferroni correction. All experiments were performed in triplicates (n=3))
A decades-old mystery in thrombosis research is solved: we present the first unified structural model of ADAMTS13 global latency, revealing its enigmatic long-range regulation is far more complex than anticipated, involving a molecular “zipper” mechanism and a pseudosubstrate role of the L3 linker.
Norman's paper on "Wrapping it Up: Structural Basis of ADAMTS13 Global Latency" is published in JTH. Congratulations, Norman!